- Messages
- 10
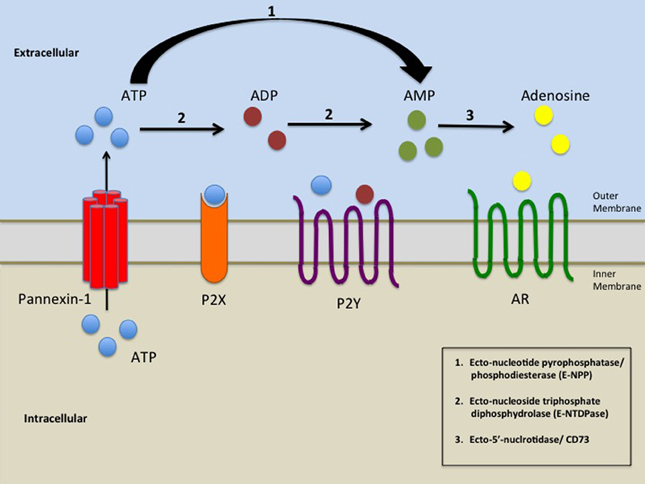
TL;DR - is it possible that instead of insufficient ATP, we're wasting it via different mechanisms than mitochondrial ATP, such as too much glycolysis?
As an (on-hold) national-level athlete, the worst symptom for me is myopathy. Most of my research centers around mechanisms to improve myopathy and fatigue, primarily skeletal muscle-related as well as general. In addition to all my own health-related research, I do a lot of tutoring in AP/college level bio, among other things, so I'm well-versed in the cellular mechanisms and energetics. I came across this today:
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5065105/
https://en.wikipedia.org/wiki/Inborn_errors_of_carbohydrate_metabolism#Galactose
Which surprised me, since everything else I've read to date seems to think that we have errors in cellular/mitochondrial energetics, sometimes specified as Complex IV/V energy transport issues. To be clear, our errors are less likely to be inborn, but we still may have issues with how our bodies process carbohydrates, which could also explain why some people do better on low carb/keto diets. As an ethical vegetarian, I did go low carb/keto for a few months, but I didn't gain much benefit, but my body did seem to appreciate going back to carbs once I discovered that ketosis is bad for muscle-fiber type switching (I need to stay fast-twitch dominant).
What really resonated with me was the one that referred to potential crossovers with severe asthma, which I also have. This makes it sound like our little hamsters are spinning in their wheels; as opposed to being unable to produce sufficient ATP, they're wasting it through too much glycolysis, which is known to speed up aging and aging-related issues.
Anyone have any insight? To the best of my knowledge, this hasn't been discussed much in these communities.
As an (on-hold) national-level athlete, the worst symptom for me is myopathy. Most of my research centers around mechanisms to improve myopathy and fatigue, primarily skeletal muscle-related as well as general. In addition to all my own health-related research, I do a lot of tutoring in AP/college level bio, among other things, so I'm well-versed in the cellular mechanisms and energetics. I came across this today:
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5065105/
https://en.wikipedia.org/wiki/Inborn_errors_of_carbohydrate_metabolism#Galactose
Which surprised me, since everything else I've read to date seems to think that we have errors in cellular/mitochondrial energetics, sometimes specified as Complex IV/V energy transport issues. To be clear, our errors are less likely to be inborn, but we still may have issues with how our bodies process carbohydrates, which could also explain why some people do better on low carb/keto diets. As an ethical vegetarian, I did go low carb/keto for a few months, but I didn't gain much benefit, but my body did seem to appreciate going back to carbs once I discovered that ketosis is bad for muscle-fiber type switching (I need to stay fast-twitch dominant).
What really resonated with me was the one that referred to potential crossovers with severe asthma, which I also have. This makes it sound like our little hamsters are spinning in their wheels; as opposed to being unable to produce sufficient ATP, they're wasting it through too much glycolysis, which is known to speed up aging and aging-related issues.
Anyone have any insight? To the best of my knowledge, this hasn't been discussed much in these communities.